

-
Review Article
February 28, 2024
English
- Exercise and adults with hemophilia: a systematic review and meta-analysis
- Doo Young Kim, Mi Yang Jeon, Young Eun, et al.
- J Korean Biol Nurs Sci. 2024;26(1):1-15.
Review Article
February 28, 2024
English
Original Article
February 23, 2024
English
Original Article
February 14, 2024
English
Original Article
February 27, 2024
English
Original Article
February 21, 2024
English
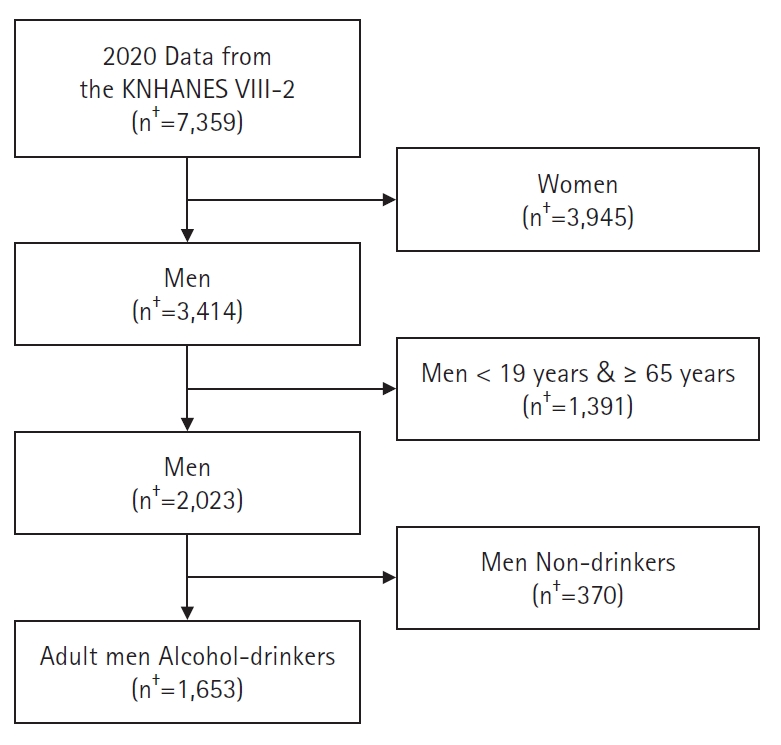
Original Article
February 26, 2024
English
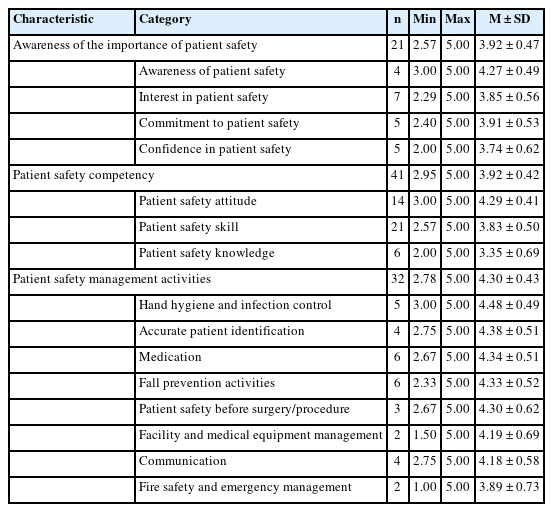

Journal of Korean
Biological
Nursing Science
Print ISSN: 2383-6415
Online ISSN: 2383-6423













Copyright © Korean Society of Biological Nursing Science.
