Morus Nigra Extract Attenuates Cognition Impairment and GABAergic Interneuron Degeneration in Aged Rat Brain
Article information
Abstract
Purpose: Aging process comes with cognitive impairment due to decreased neuronal cell number, activity, and neuronal circuit. Alteration of inhibitory neurons contributes to cognitive impairment in normal aging and is responsible for disrupting the excitation/inhibition balance by reducing the synthesis of gamma-aminobutyric acid (GABA). Morus nigra (Mulberry) is a natural physiologically active substance that has been proven to have anti-oxidant, anti-diabetic, and anti-inflammatory effects through many studies. This study aimed to evaluate the effects of the mulberry extract (ME) on cognitive function through anti-oxidant enzyme and GABAergic neuronal activity in aged rat brain. Methods: Sprague Dawley rats were randomly assigned as the young group (8 weeks, n=8), aging group (67 weeks, n=8), and aging+mulberry extract group (67 weeks, n=8). The aging+mulberry extract group was orally administered 500 mg/kg/d mulberry extract for 6 weeks. Results: The aging+mulberry extract group improved spatial and short-term memory. The antioxidant potential of ME increased the expression of superoxide dismutase-1 (SOD-1) and decreased inducible nitric oxide synthase (iNOS). Also, the aging+mulberry extract group significantly increased the expression of GABAergic interneuron in hippocampus cornu ammonis1 (CA1) compared to the aging group. Conclusion: The number of GABAergic inhibitory interneurons was deceased and memory functions in the aging process, but those symptoms were improved and restored by mulberry extract administration.
INTRODUCTION
Aging is inevitable for all organisms, even without a pathological condition. Normal aging progressively worsens with age, making irreversible changes in all physiological processes. It affects the homeostasis of the endocrine system and the heart, lungs, liver, bladder, muscles, and skin [1]. Of the many changes, aging is the most significant influence on cognitive decline in brain function. The primary cognitive function most affected by age is attention and memory, including perception, thinking, reasoning, memory, analysis, concentration, judgment, and insight [2]. The hippocampus, part of the brain related to cognitive function, plays an essential role in learning and memory. Since the hippocampus is directly involved in pathfinding, episode experience, working memory, and attention, hippocampal damage affects spatial memory and short-term memory. The cause of aging has been argued for a long time, and oxidative stress is one of the most influential culprits. The overproduction of reactive oxidative species (ROS) produced by mitochondrial respiration is highly involved in oxidative stress. Most ROS within mitochondria eventually damages membrane phospholipids, intracellular proteins, and DNA, offering them to be impaired predominantly neuronal cells in the brain. Increased oxidative stress and a concomitant decrease in the brain’s anti-oxidant defense system have been associated with age-related cognitive decline [3]. In general, aging increases free radicals and neuroinflammatory responses. Long-lasting pro-inflammatory cytokines produce memory dysfunction, especially in the hippocampus [4].
Gamma-aminobutyric acid (GABA), a neurotransmitter, accounts for 30% of all neurotransmitters in the central nervous system and is present at high concentrations. GABA, an inhibitory neurotransmitter, effectively regulates neuronal stabilization and improves memory by arbitration of neuronal excitability and improving brain metabolism and blood flow [5]. Difficulty in regulating GABA and glutamate occurs even in normal aging. The inhibition imbalance may be due to increased glutamate (excited) signal transduction or reduced inhibition due to decreased GABAergic signal transduction [6]. GABAergic interneurons play essential roles in neural circuitry and activity. Inhibitory neurons represent the most complex and diverse group of neurons and possess a solid ability to control physiological responses [7]. Inhibitory neurons play an important role in regulating cortical function and plasticity, and aging can impair this function [8].
Mulberry is a mature fruit of the Moraceae tree, and its scientific name is Morus. The black color of mulberry fruits contains anthocyanins and has a relatively higher content than other fruits. Anthocyanins are flavonoids, antifoam, anti-allergic, immune biologically active agents such as antiviral agents were confirmed [9]. The anthocyanins of the mulberry are the components of cyanidin-3-glucoside and cyanidin-3-rutinoside, which has a scavenging function of hydrogen peroxide, a free radical [10]. Mulberry also contains natural GABA, among many bioactive compounds, and mulberry leaf tea with GABA may help control blood pressure, blood lipids, and blood sugar [11]. However, no studies have reported mulberry’s functionality on GABAergic interneurons associated with brain aging.
Therefore, this study aims to investigate the cognitive function of aged rats treated with mulberry extract and examine the neurological changes of the GABAergic interneuron.
METHODS
1. Animals & Experimental design
Sprague-Dawley (SD) male rats (8 weeks & 67 weeks old, Orient. Bio, Korea) were randomly assigned to each group after 1 week of adaptation. Because SD rats have a higher incidence of kidney and brain tumors at 2 years, we investigated the normal aging process using an 18-month-old. During the experimental period, male SD rats were given appropriate conditions to live in animals (temperature 23±2°C, humidity 50±5%, and 12 hours lighting cycles). As an experimental design, rats were randomly grouped: group 1: young control group (Y-CON, n=8); group 2: aging control group (A-CON, n=8); group 3: aging+mulberry extract group (A-ME, n=8). After acclimation for 7 days, the experimental group was administered mulberry extract (500 mg/kg/d) with gavage at the same time every day for 6 weeks. And then, two types of behavioral tests were performed, and rats were sacrificed the next day without pain.
2. Preparation of mulberry extract (ME)
Dried mulberry fruit 300 g is purchased (Buan, Korea) and extracted into 1 L of 80% ethanol solution, and the active ingredient is extracted for one day. The extract was then filtered on Whatman No.3 filter paper. Thereafter, the filtrate was concentrated under reduced pressure, and the ethanol of the purified extract was evaporated to obtain a mulberry extract.
3. Brain preparation
After all, experiments were completed, rats were placed in a carbon dioxide (CO2) chamber and under anesthesia by inhalation. After visual observation of respiratory arrest, the gas flow was maintained for at least 1 minute, the chest cavity was opened under deep anesthesia, a 24 G needle was inserted into the heart’s left ventricle, and 0.05 M phosphate-buffered saline (PBS) was inserted. After the blood was removed entirely, using a solution of 4% paraformaldehyde (PFA) in the same ratio, performed at a constant rate of 50-60 mL/min for 10 minutes to fix the tissue. The extracted brain is further fixed with 4% PFA solution at 4°C for 24 hours. After that, it was precipitated in a 30% sucrose solution for 2 to 5 days to prevent crystallization in the tissue and cut into 40 μm slices using a tissue cutting machine Cryostat (LeicaCM3050S, Leica, Berlin, Germany), and immunohistochemistry was performed. For Western blotting, all blood was removed by perfusion with 0.05 M PBS, and then brains were stored frozen at -70°C immediately.
4. Immunohistochemistry
The hippocampal sections of each group were washed three times with 0.05 M PBS for 5 minutes each and then 3% H2O2 for 30 minutes to remove endogenous peroxidase. After that, wash with 0.05 M PBS until bubbles disappear and react with a blocking solution for 2 hours. The primary antibodies: anti-Parvalbumin (1:500, Abcam, AB11427), anti-Calbindin (1:500, Abcam, AB11426) were incubated overnight at 4°C. The secondary antibody: anti-Goat rabbit (1:500, Santacruz, SC02004, CA, USA), was reacted at room temperature for 1 hour. The sections were washed for 5 minutes in 0.05 M PBS and reacted with avidin-biotin complex (ABC, Vectastain-Elite ABC kit, Vector, Burlingame, CA, USA) for 1 hour. After washing, the sections were stained with diaminobenzidine (DAB Vector, Burlingame, CA, USA) and washed again with 0.05 M PBS. And then, the sections were attached to the gelatin-coated slide glass by the free-floating method and dried naturally overnight at room temperature. After drying, the sections were dehydrated with 70%, 80%, 90%, and 100% ethanol and were made transparent with xylene. The cover slides are covered using a permount (Fisher Scientific, Pittsburgh, PA). Finally, positive cells were observed using an optical microscope (BX51, Olympus, Tokyo, Japan) of 100×, 400× magnification.
5. Western blot analysis
Proteins were extracted from hippocampal brain tissue using a low-temperature radioimmunoprecipitation assay (RIPA) lysis buffer (EBA-1149, ELPIS Biotechnology, Daejeon, Korea) containing a protease inhibition cocktail (Thermo Fisher Scientific) and a phosphate inhibition cocktail (Cell Signaling Technology). Used a machine to homogenize and then centrifuge (Centrifuge 5415R, 13,000 rpm, 20 minutes, 4°C) to obtain the supernatant. The supernatant was quantified at 5-10 μg and electrophoresis at 8-12% sodium dodecyl sulfate-polyacrylic amide (SDS-PAGE) gel (30V for 0.5 hours and then 100V for 2 hours). After electrophoresis, the protein is transferred to a polyvinylidene fluoride (PVDF) membrane (Merck Millipore, ISEQ00010). The membrane was blocked with 5% skim milk for 1 hour at room temperature. With the following primary antibodies: anti-Parvalbumin (1:1,000, Abcam, AB11427, USA), anti-glutamic acid decarboxylase (GAD) 67 (1:2,000, Abcam, AB26116, USA), anti-superoxide dismutase-1 (SOD-1) (1:1,000, Santa Cruz 4, SC1407, USA), anti- glutathione transferase omega 1/2 (GSTO1/2) (1:1,000, Santa Cruz, SC98560, USA), and anti-β-actin (1: 15,000, Santa Cruz 4, SC47778, USA) was reacted for overnight at 4°C. The membrane was washed three times with tris-buffered saline with 0.1% Tween® 20 detergent (TBST) and reacted with horseradish peroxidase (HRP)-conjugated secondary antibody for 1 hour at room temperature: anti-rabbit antibody (1:5,000) or anti-mouse antibody (1:5,000). And then, washing with TBST, the protein quantity was visualized with enhanced chemiluminescence (ECL) to detect the emitted light. Finally, development was carried out on an X-ray film (Agfa, Mortsel, Belgium), and density was measured in pixels using Image J software (U.S. National Institutes of Health, Maryland, USA). Each measured value is quantified based on β-actin.
6. Cognitive function tests
1) Morris water maze test
The Morris water maze test (MWM) is a test for evaluating learning and memory [12]. Prepare around the pool with a diameter of 200 cm and a height of 35 cm, a platform with a diameter of 15 cm, and a Video Camera (S-MART: Pan-lab, Barcelona, Spain) connected to the tracking device. Fill water with a temperature of 22±2°C to the height of 2 cm above the platform. Then, the water in the pool was made black using harmless paints, making the hidden platform invisible. Provide a visual marker to remember the location of the platform. The procedure of this test was trials per day for 4 consecutive days, and, after that, a probe trial was performed on day 5 for each rat. Rats are placed in one of four pool quadrants to find the hidden platform. The quadrant is repositioned to minimize the possibility of accidentally going to the platform by varying the location of each place in the pool. The time required for the rat to go to the platform was defined as the mean escape latency. The mean value of the escape latency time was obtained three times a day. The maximum test time was set to 60 seconds.
2) Passive avoidance test
The passive avoidance test identifies short-term memory for the aversion stimulus (electricity) [13]. Gemini (San Diego Instrument, San Diego, CA, USA) used the equipment equipped with an automated system. The box is divided into a bright light hate space and a dark, mysterious space, and the floor of the shelter room can be electrical stimulation. There is a door that automatically closes between the two areas. The experimental method is as follows. When a rat is placed in a bright light hate space on the first day, the rats instinctively avoid a dark area. The door is closed by the sensor, and the foot shock (0.3 mA of electric shock) is applied for 2 seconds. The second day is 24 hours after the first day. Proceed as in the first experiment but do not give a shock. On the second day, record the time of avoidance from the bright to the dark space, even if there is no electric shock, as the latency time. The time limit is set to 300 seconds, and when the time limit is exceeded, the time is set to 300 seconds.
7. Research Ethics
This study was approved by Institutional Animal Care and Use Committee (IACUC) and K University Institutional Animal Care Committee (KHUASP-170-010). The handling of laboratory animals and all laboratory procedures are described in the NIH Guide for the Care and Use of Laboratory Animals. All the experimental procedures were performed following the National Institutes of Health (NIH).
8. Statistical analysis
Statistical analysis the results of this study were analyzed using SPSS version 23.0. Data are presented as mean±SEM. Comparisons among groups were performed by the One-way analysis of variance (ANOVA). Post hoc was done by Scheffe’s test for comparison between the three groups. Differences are considered significant to be p-value <.05.
RESULTS
1. Effect of ME on the cognitive function in the aged rat
To confirm the enhancement effect of spatial memory, we conducted an MWM test. From the first day of training, there was a significant difference in that the aging group was delayed compared to the Y-CON group (F=11.30, p<.001). On the fourth day, the Y-CON group recorded, 23.51±1.89 seconds, the A-CON group recorded 53.27±2.18 seconds, and the A-ME group recorded 26.59±1.72 seconds (F=38.59, p<.001). There was a significant difference between the A-ME and A-CON groups. The A-ME group improved spatial memory similar to Y-CON on day 4 (Figure 1-A). Passive avoidance experiments were performed to assess the degree of short-term memory impairment due to aging. The latency times of the Y-CON group were highest at 300 seconds, the A-CON group was 130.00±45.79 seconds, and the A-ME group was recorded at 281.43±18.57 seconds. After Scheffe’s post-analysis, latency was significantly reduced in the A-CON group, and the A-ME group was no different from the Y-CON group (F=10.68, p<.001). These results showed that the A-ME group alleviates short-term memory corruption more than the A-CON group (Figure 1-B).

Effect of mulberry extracts on cognitive function. (1-A) Morris water maze test, (1-B) Passive avoidance test. *p<.05, significantly different from Y-CON. #p<.05, significantly different from A-CON. Y-CON=Young control group; A-CON=Aging control group; A-ME=Aging mulberry extract group.
2. Effect of ME on antioxidant enzymes in the aged rat hippocampus
To determine the antioxidant enzyme activity associated with aging, the protein expression of SOD-1 and GSTO1/2 in the brain’s hippocampus was quantitatively analyzed (Figure 2). The relative optical density in SOD-1 expression levels was significantly decreased to 0.78±0.04 in aging rats compared to young rats. The experimental group of aging rats treated with ME was 0.90±0.01, significantly higher than that of the aging rat control group and smaller than that of the young rat control group (F=41.17, p<.001). The relative optical density in GSTO1/2 expression levels was not significantly different in the 3 groups (F=0.90, p=.429).
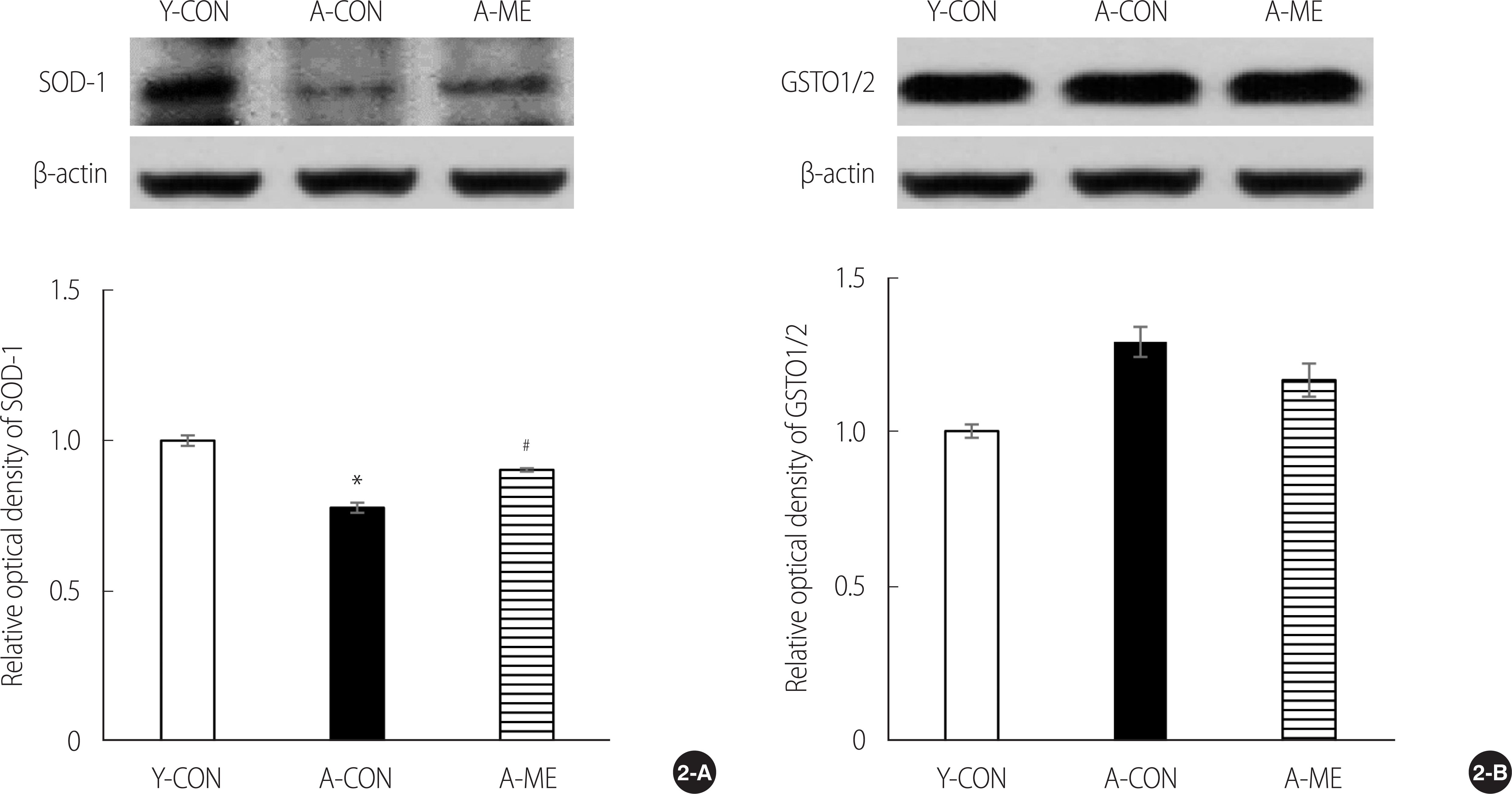
Western blot analysis and quantification of anti-oxidant enzymes in the hippocampus. (2-A) The expression of SOD-1, (2-B) The expression of GSTO1/2. Data are presented as the mean±SEM. *p<.05, significantly different from Y-CON. #p<.05, significantly different from A-CON. Y-CON=Young control group; A-CON=Aging control group; A-ME=Aging mulberry extract group; SOD-1=Superoxide dismutase-1; GSTO=Glutathione s-transferase omega.
3. Effect of ME on inflammation in the aged rat hippocampus
To confirm ME treatment on inflammation response in hippocampal tissues, iNOS expression was confirmed by western blot. As a result, the A-CON expression of iNOS was higher than that of Y-CON and significantly decreased in the treated A-ME group (F=29.38, p<.001) (Figure 3).
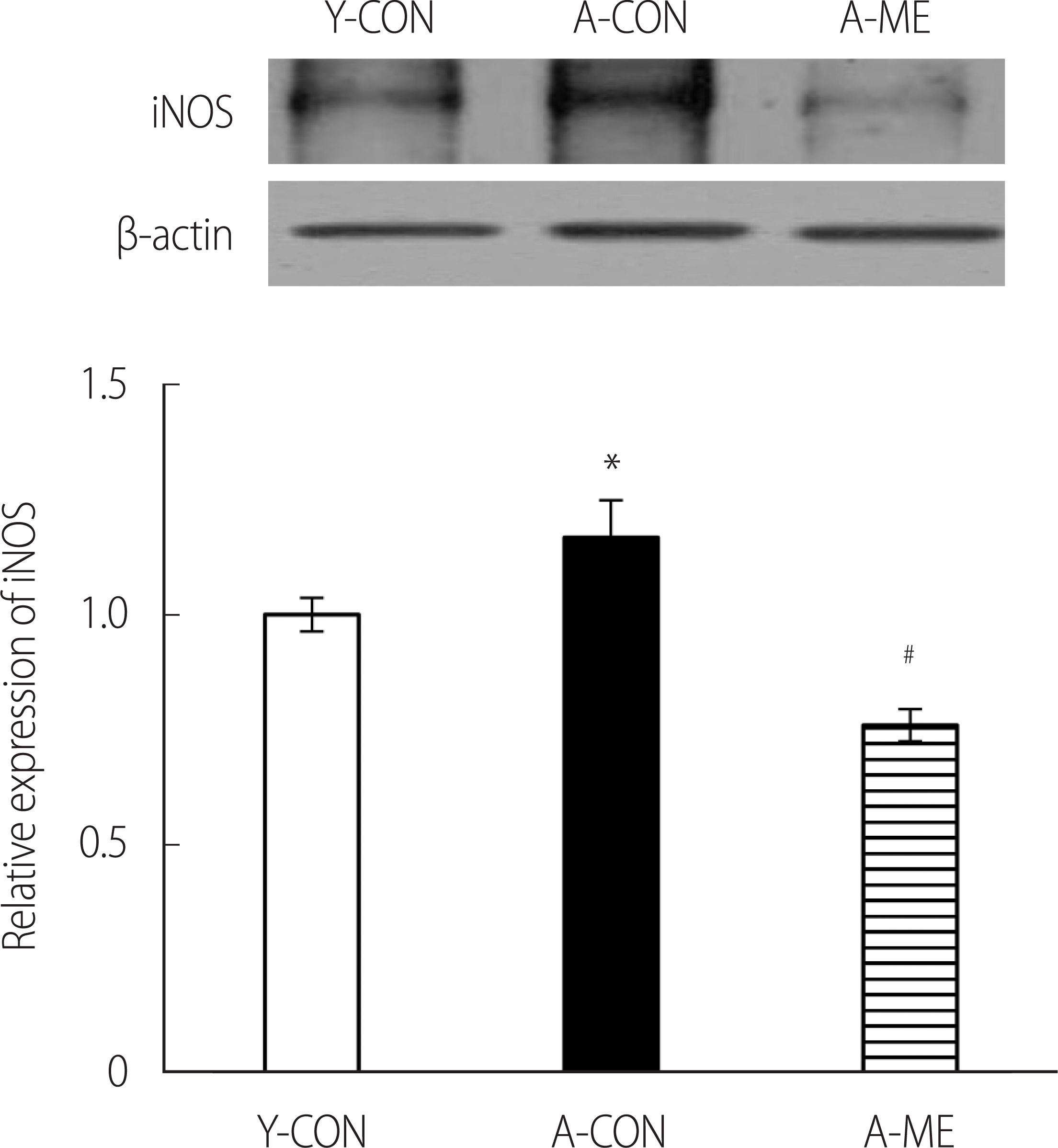
Western blot analysis and quantification of iNOS in the hippocampus. Data are presented as the mean±SEM. *p<.05, significantly different from Y-CON. #p<.05, significantly different from A-CON. Y-CON=Young control group; A-CON=Aging control group; A-ME=Aging mulberry extract group; iNOS=Inducible nitric oxide synthase.
4. Effect of ME in NeuN positive cell in the aged rat motor cortex
The number of positive cells was counted by selecting the grid size (150×150 μm) from each group of the motor cortex. The well-known neuronal marker, neuron-specific nucleoprotein (NeuN) showed a significant decrease in the number of neurons in aging, with 140.20±2.70 in Y-CON and 119.80±1.40 in A-CON (F=16.18, p<.001). The positive cell number of the A-ME group was 128.30±3.50, which was higher than that of the A-CON group, but there was no significant difference (p=.098) (Figure 4).

The number of NeuN positive cells in motor cortex. Data are presented as the mean±SEM. *p<.05, significantly different from Y-CON. #p<.05, significantly different from A-CON. Y-CON=Young control group; A-CON=Aging control group; A-ME=Aging mulberry extract group; NeuN=Neuron-specific nucleoprotein.
5. Effect of ME on the decrease of GABAergic interneuron in aged rat hippocampus
To identify GABAergic interneurons related to cognitive function in the brain’s hippocampus, GAD67, calcium-binding proteins parvalbumin, and calbindin were determined by western blot and immunohistochemistry (Figure 5). GAD67 expression levels greatly reduced A-CON and increased from A-ME to Y-CON level (F=23.31, p<.001) (Figure 5-C). Parvalbumin (PV) also had the same result (F=23.71, p<.001) (Figure 5-A). The number of GABAergic immuno-positive neurons was measured in the CA1 layer. Results of immunohistochemistry in PV showed a significant decrease in the effective neuron of A-CON compared to Y-CON and restoration to the Y-CON level at A-ME (F=21.79, p<.001) (Figure 5-B). The same result was found in calbindin (F=46.17, p<.001) (Figure 5-D).
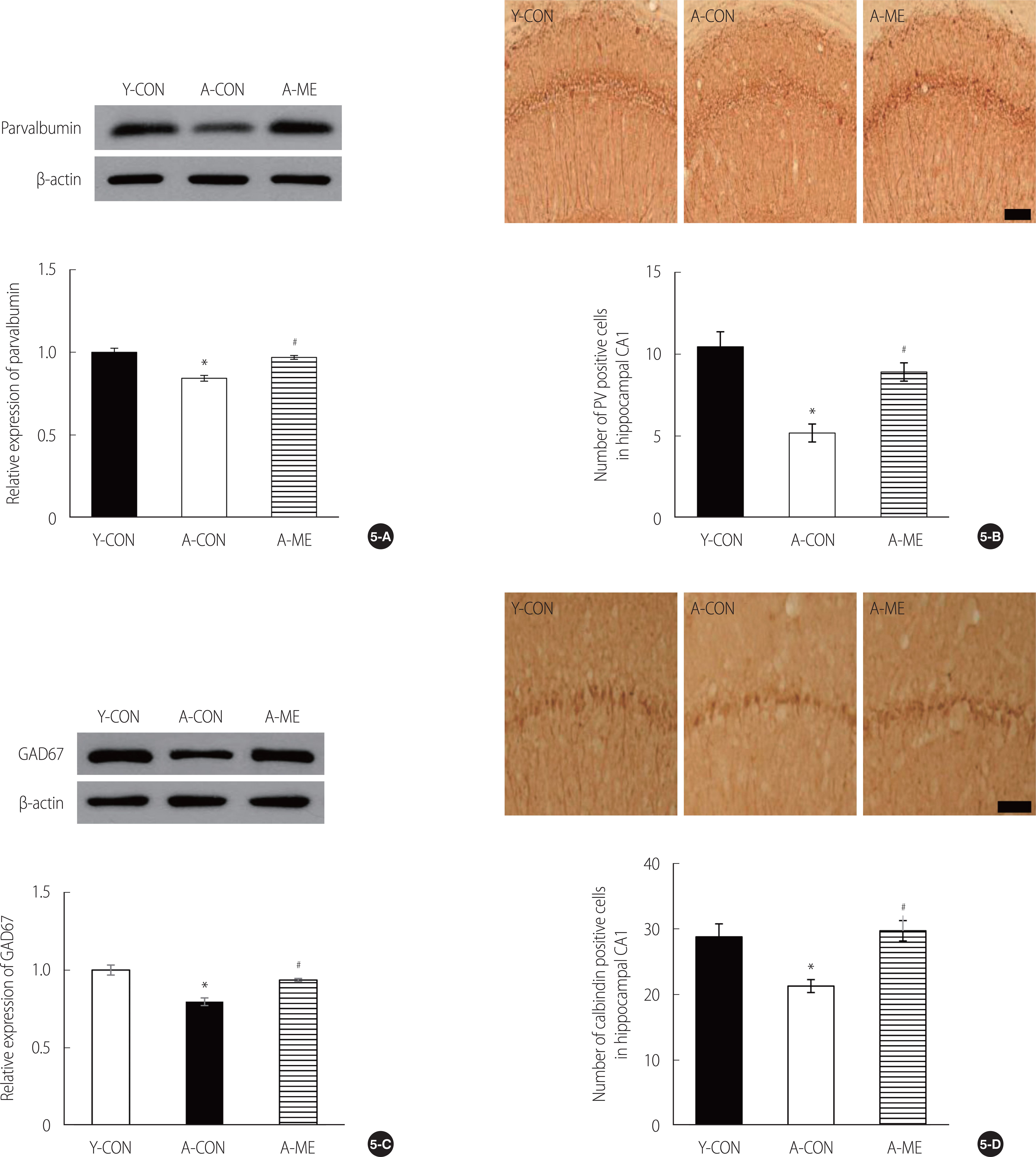
The result of Gabanergic neuron in in aged rat hippocampus. (5-A) The expression level of Parvalbumin, (5-B) The number of Parvalbumin positive cells in the hippocampal CA1 layer, (5-C) The expression of GAD67, (5-D) The number of Calbindin positive cells in the hippocampal CA1 layer. Data are presented as the mean±SEM *significantly different from Y-CON. #significantly different from A-CON. Y-CON=Young control group; A-CON=Aging control group; A-ME=Aging mulberry extract group; CA1=Cornu ammonis 1; GAD=Glutamic acid decarboxylase; PV=Parvalbumin.
DISCUSSION
This study aims to investigate the effect of natural mulberry fruit extract on GABAergic neurons in aged rats’ brains. As a result, oral administration of mulberry extract effectively improved the cognitive decline of aged rats by reducing the dysfunction of inhibitory neurons during normal aging. Understanding normal brain aging is essential for finding ways to overcome and maintain cognitive dysfunction associated with aging.
Aging is characterized by chronic inflammation called senoinflammation. Alleviating inflammation plays an important role in diseases and conditions associated with aging [14]. Previous studies show that mulberry’s dark red and black color contains anthocyanins, and it has been reported to be effective in inhibiting inflammation and removing ROS scavenging [10]. Our data confirmed the restoration of SOD-1 in the mulberry administrated aging group. In addition, it was confirmed that the activity of an inflammation-inducing factor, iNOS, was inhibited. SOD is a universal enzyme in organisms that live in the presence of oxygen [15]. SOD is the only anti-oxidant enzyme that directly removes ROS and the first line of defense mechanism against superoxide free radicals, catalyzing peroxide conversion into oxygen and hydrogen peroxide [16]. Glutathione transferase Omega 1,2 is the most recently defined class of glutathione S-transferases (GSTs) and plays a vital role in reducing oxidative stress produced by various stressors and cellular processes [17]. GSTO has protection against neuron damage, but the molecular mechanism and physiological functions of GSTO are still unclear, and further research is needed [18]. In this study, the activity of SOD-1 was significantly recovered in the mulberry treated aging group, but there was no statistically significant difference in GSTO1/2. It may be due to the competition of the same enzyme mechanism.
The aging-related functional decline of the brain leads to disarrangement of inhibitory neurotransmission, which can be confirmed by GABAergic neuronal expression. GAD, a GABA synthetase, is available in two forms, molecular weight 67 kDa or 65 kDa, present in the brain. GAD65 is confined to the axon terminal in synapses, while GAD67 plays a role in protecting and generating synapses at neuronal injury [19]. In physiological or pathological processes, GAD67 modulates the activity of neurons. GAD67 expression reflects a circuit activity level that continuously regulates the inhibition intensity by increasing the suppression output at high circuit activity and decreasing the output at the low activity [20]. PV is approximately 25% of primate dorsolateral prefrontal cortex GABAergic neurons [21] in pathological conditions and is expressed in the chandelier, basket cells, and Purkinje cell layers. PV is known to control spike timing in adjacent excitatory neurons. Also, PV is strongly involved in intracellular calcium regulation and trafficking, which binds calcium with high affinity. The maintenance of calcium homeostasis is also important in brain aging. Various oxidants cause calcium influx and affect mitochondria and nuclei. As a result, calcium is used as a signaling molecule in response to the stimulation of oxidants [22]. Calcium-binding protein (CBP) binds Ca2+ ions and regulates Ca2+ concentration in the neuronal cytoplasm. CBP does not act as a direct neurotransmitter but as a cellular function signal and protects neurons by preventing excessive calcium ion influx during membrane potential excitation [23,24]. The overall homeostasis of the nervous system is maintained by maintaining a regulated rate of network function of excitatory or inhibitory neurons [25]. The synaptic imbalance between excitation and inhibition (low E/I ratio or high E/I ratio) in the cortex can impair the function of decision making, a key factor in cognitive function [26]. In addition, E/I imbalance can underlie the pathology of aging and neurodegenerative diseases [27]. We identified disturbances in E/I balance as inhibitory neuronal GABAergic deficits but not excitatory imbalances. However, excitatory imbalances often result from decreased GABA A receptor activity [27]. Nevertheless, oxidative stress dysfunction has been reported to be exacerbated by hyperexcitation [25], so it is necessary to investigate excitatory neuropathology in future studies.
This study shows that aging reduces the expression of GAD67 in the hippocampus, which is consistent with previous findings. Also, it was confirmed that the expression of PV and Calbindin also decreased in aging [28]. The expression levels of GAD67 and CBP were increased in the experimental group to which the mulberry extract was orally administered. Interestingly, there was no significant difference in the expression of NeuN, a neuronal marker of the brain. However, further studies are needed to determine whether ME is specific for GABAergic neurons.
This study found that the administration of the mulberry extract restored the reduction of inhibitory GABAergic intermediate neurons due to aging, and cognitive impairment was significantly improved. Most previous studies only revealed differences between ages and animal types. Also, the study of mulberry reported studies focused on anti-oxidant properties. However, the present study investigated the degree of cognitive recovery after oral administration of physiologically active mulberry extract in normal aging rats by altering the immune responses of GABAergic interneurons and calcium-binding proteins. And this study may help to understand the effects of anti-oxidant therapy on age-related cognitive impairment.
CONCLUSION
Cognitive decline associated with aging arises from the dysfunction of inhibitory GABAergic interneurons. It is well known that excessive free radical production, or failure of free radical scavenging, and chronic inflammation act as nervous system toxicity during the aging process. This study indicated that such neurotoxicity can be protected by oral administration of mulberry extract rich in physiologically active substances. This study on the deterioration of cognitive function accompanied by the general aging process is expected to be useful for future studies of aging-related diseases. However, further studies are needed on the duration of administration, the concentration of administration, and the oxidative potential of the individual. Also, Further research is required to explore molecular mechanisms to improve the therapeutic effect of the active ingredient. It is suggested that the study of each component’s brain response and efficacy is required.
CONFLICT OF INTEREST
The authors declared no conflict of interest.
AUTHORSHIP
JHL and YJK contributed to the conception and design of this study; JHL, YJK, and MKS collected data; JHL performed the statistical analysis and interpretation; JHL drafted the manuscript; JHL and YJK critically revised the manuscript; YJK supervised the whole study process. All authors read and approved the final manuscript.